开源学是什么
后天规则分析工具概述
开源学是什么
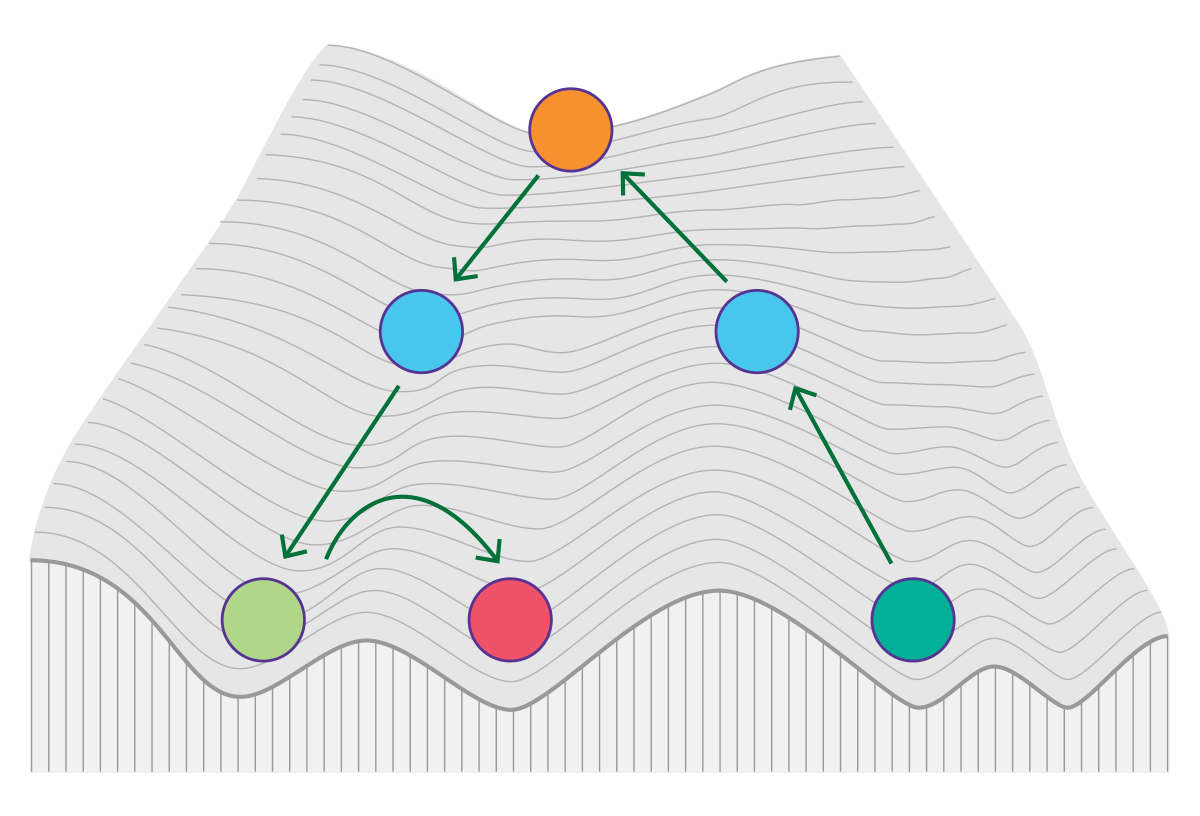
遗传学研究基因表达方式变化由机制而非底层脱氧核糖核酸序列变化引起的可这到底意味着什么
所有多细胞生物(象你!)都包含核内大致相同的DNA序列,遍及所有细胞类型,但皮肤细胞可与肝细胞和心机等有独特区别。机制建立这些特殊细胞类型 被首先确认为
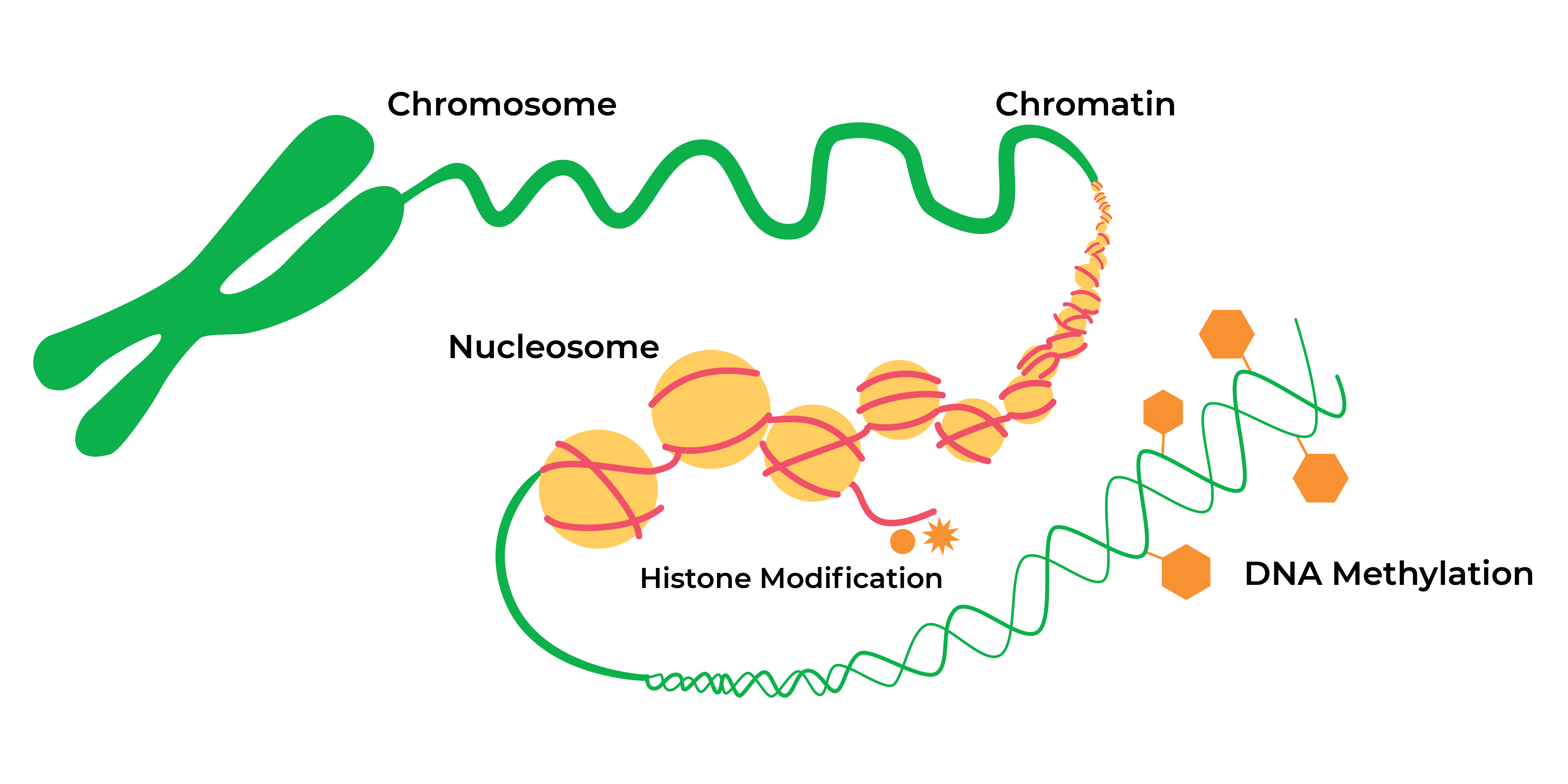
希腊前缀epi-表示Nations序列本身历经数十年研究后变异或标记,现今变异泛指可遗传直通修改和脱氧核糖核酸脱甲基化而不改变可影响基因活动底层脱氧核糖核酸序列
后代调节面大为扩展histone修改法(如吸附和甘油残留核蛋白甲基化法)有助于扩展或压缩染色素成开机式或闭机式配置与开机或离机式基因表达法相关表示某些非编码RNA帮助规范转录过程或之后的基因表达脱氧核糖核酸甲基化是另一个关键修改法,即跨基因组CpG内嵌入细胞序列
后天变换函数是什么

所有这些不同标志的最终功能是协调调节基因表达研究显示这些标记可起分子机制作用 介导环境对基因组的影响 从胚胎开发开始举例说,母亲早孕期间的生活方式和环境条件可能对子孙的健康产生长期影响。一号脱氧核糖核酸ameome报告受母体吸烟影响2事实上,有新证据显示,有些后代标志代代相传,尽管这些标志多半先在胚胎早期消除出生时和我们老化时 脱氧脱氧模式随时间变化最近的研究还发现,特定地缘的甲基化水平可用作人生物时代预测器3
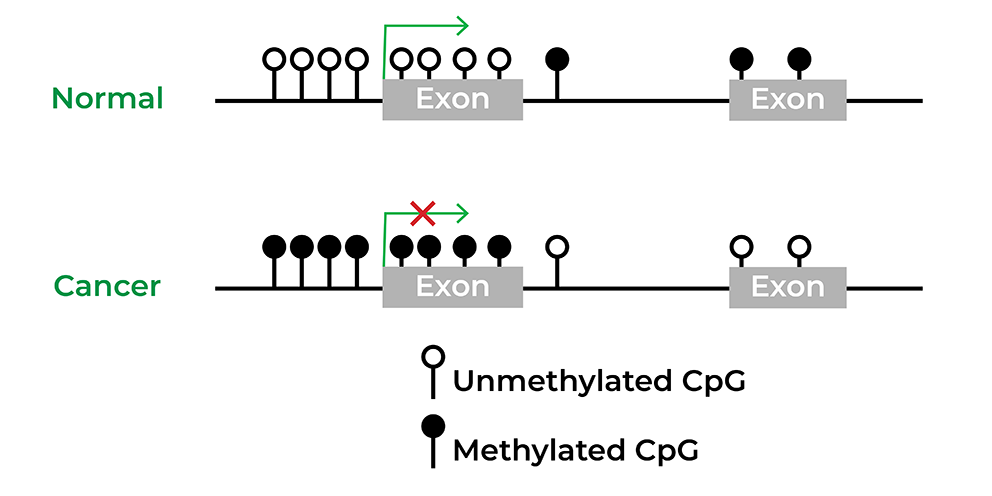
遗传学还广泛与疾病相关联,特别是癌症数项创举如癌症基因组图集或TCGA快速开发以获取人类失序特征利用大群研究发现生物标志,甚至商业化从结肠癌到膀胱癌等各种癌症4突变药证明有效处理血科肿瘤和固态肿瘤,许多临床试验正在进行中以开发单面向癌症上位或与其他理疗相联的小模块抑制器5
如何研究后代变换
最特征化后代标志脱氧核糖核酸.标准排序方法无法直接分析脱氧脱氧法,因为Metro组与Citosine(5-Metrocetsine)有共价绑定5-mC)Bisulfite转换视黄金标准识别5-mC变化化学过程解析细胞素并转换成uracil使用NGS变位法算法算法算法为C类,转成uracil的C类则算法为T类通过比较这些序列和参考基因组,人们可以计算单基解析法的甲基化百分比减少表示双词顺序,全Genomebisulfite定序并目标二叉线程最疯狂使用NGS技术剖析methome
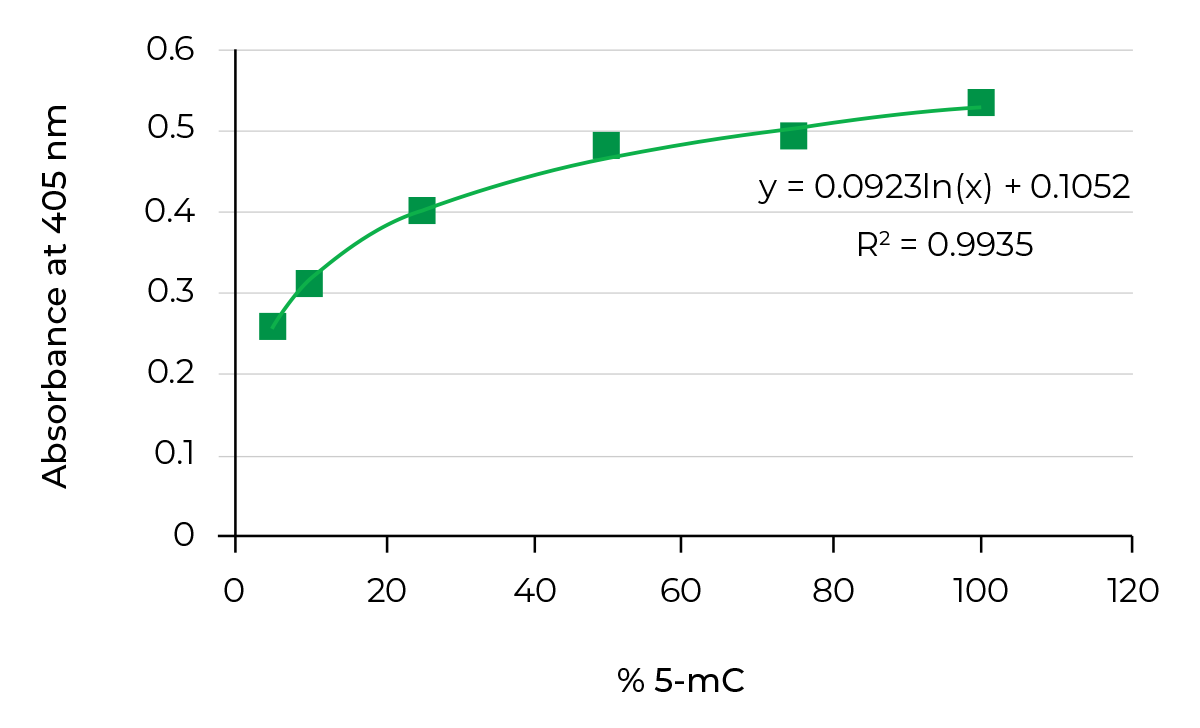
反之,无二叉法,如MateddDNA免疫降水MeDIP使用抗体对准5mC从零散基因组脱氧核糖核酸池中丰富脱氧丰度分片后可用于全基因组甲基化分析研究者也可以用同一种抗体对目标5mc并用ELISA技术类
如何分析chromatin结构变化
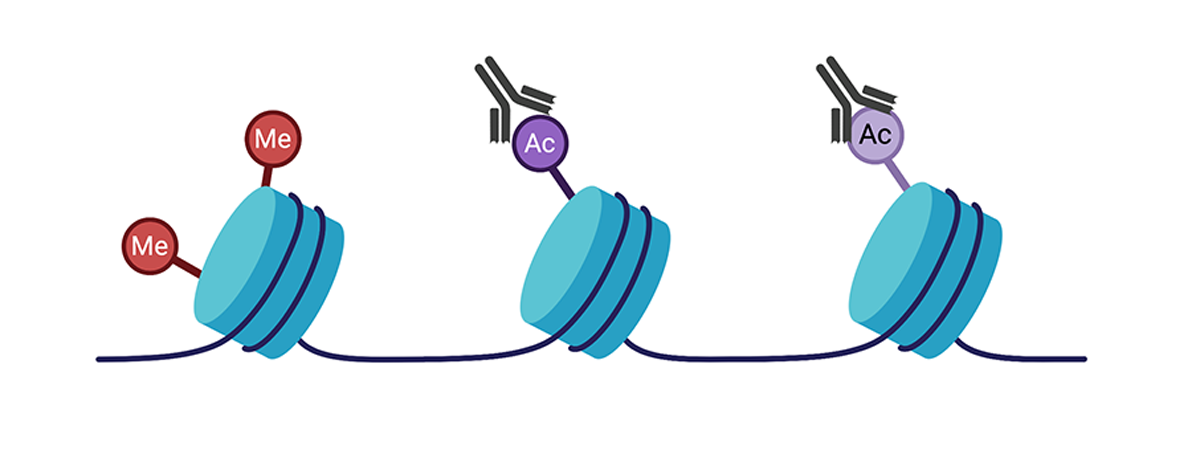
histone修改也是核心直方蛋白共价修改,这也可能改变基因表达式。蛋白质与脱氧核糖核酸交叉连接后,可使用蛋白质专用抗体选择沉浸脱氧核糖核酸碎片染色体免疫溢出.浓缩脱氧核糖核酸碎片可排序ChIPseq由下G定序实现基因组广度变换ChIP-seq方法也可以用于分析转录因子和基因组中的其他酶绑定点
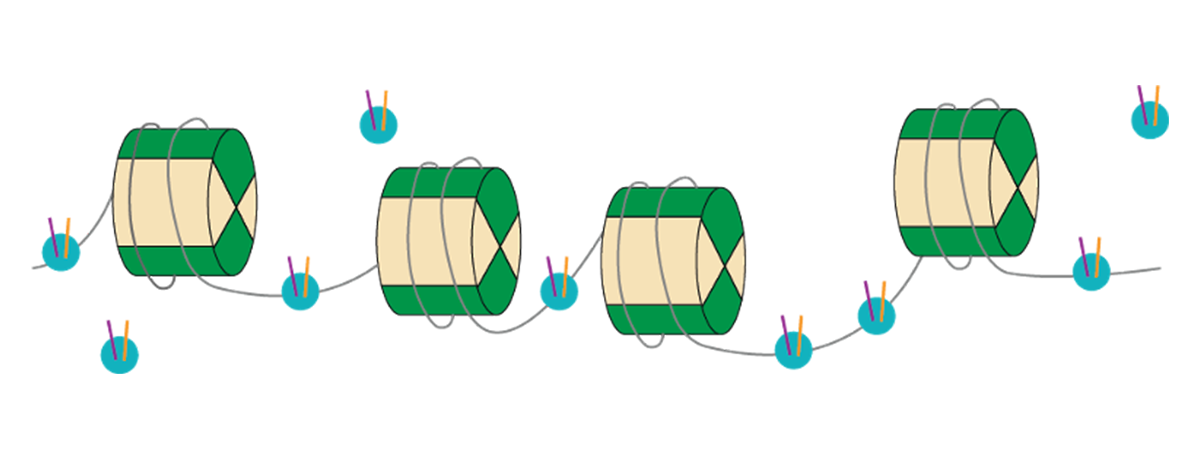
Chromatin可获取性通常与基因表达方式相关联,测量开chromatin的存在或缺失的方法帮助我们理解染色素结构改变后台景观的重要性最常用染色素无障碍技术ATAC-seq允许研究人员查看脱氧核糖核酸存取强法利用转录器只能插入适配器序列到开染色素网站深入测序覆盖范围,还有可能辨别独特的绑定网站或抄录因子脚印,这些因子可能绑定在原生染色素内部
下一个词学研究
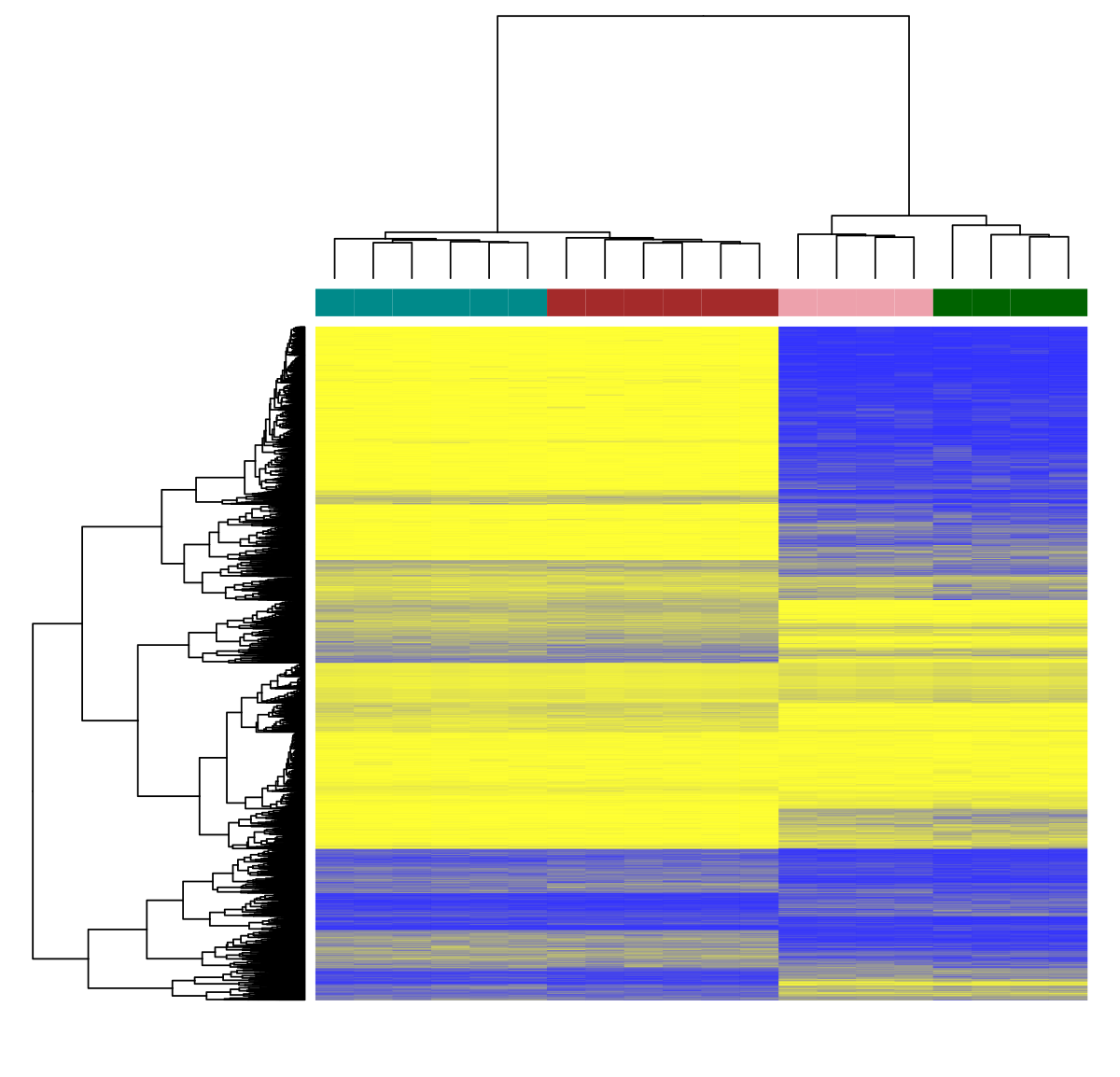
结合所有上述方法与RNA排序,现在有可能使用多组化方法直接测量后代变化的影响举例说,合并甲基化标记资料、染色素可获取性资料和基因表达方式可能揭示环境驱动的独特机制6.
更强大的是单细胞内应用这些方法最近测序电量快速增加并大幅下降成本后,对单细胞分解生成细胞型图的兴趣猛增人类细胞图集项目 由数位杰出研究者引导 在这个前沿领域 努力创建全方位参考图允许新层次理解细胞开发、特征和功能举个例子全原二叉定序实验单细胞鼠肝脏显示肝组织内异异度之高出奇7测序物价持续下降并开发新方法后,
精品指南
| 脱氧核糖核酸分析 | |
|---|---|
| 双工法 | |
| Bisulfite转换 | EZ脱氧核糖核酸 |
| NGS平台 | |
| PCR平台 | |
| Bisulfite免费方法 | |
| 反体方法 | |
| 复元方法 | |
| 染色体分析 | |
| ChIP系统 | |
| Chromatin结构 | ymo-seqATAC |
引用
- ToranoEG.加西亚市G.费尔南德斯-莫雷拉L.Nino-Garcia P费尔南德斯F.,外部因素对Epenome:Utero和Liver生化Res int2016 268635
- 朱伯特R.菲利克斯JF.优素菲P巴库尔斯基M.简单AC.布雷顿市里瑟SE.马库纳斯市..!里士满C.徐市J.Küpers L.KS. S.S.浩阳市格鲁齐耶娃Söderhäll,C.萨拉斯市..!拜兹市张Hlepeule,J.Ruiz CLigthart S.王泰泰勒J..!Duijts L.夏普GC.扬基派信..!尼尔森市M.瓦兹A法林市D.湖德利通州..!富姆梅勒F.金云基尔J库尔lMunthe-KaasC.盖林市布斯塔曼特M索尔-库比佐勒斯市J.Quraishi市M.任J托斯特JGonzalez JR.彼得斯市J.哈博格市E.徐Z范默尔斯....高特R.KerkhofMCorpelijnE芬贝格AP.EngC巴卡雷利..!本杰明尼龙E.布拉德曼A默里德市K贝格斯特伦A黑塞格ZHernandez-Vargas布鲁内克里夫皮那特M休德B以华特S姚JLemonnier,N.佛朗哥OH.吴MC.霍夫曼AMacArdleWVan derVies P法拉希F吉尔曼市W.Barcellos L.F.库马尔A威克曼MGuerra S查尔斯M..!霍洛维JAffray C铁米埃W.史密斯GD.邮报D希弗特MF.Eskenazi,B.弗列海德 M阿沙德HAntó JM.德汉A卡尔马乌斯W安西梅萨诺松叶尔J甘图斯A伯沙根GHolland N墨菲SK德米欧市L.布尔沙德G.Ladd-AcostaC斯涅德 H尼斯塔德WKoppelman GH.列尔顿市L.Jaddoe,VW.威尔库克斯MelénE伦敦SJ.DGNDMMTEDLEGEUMETAlyServe:Geneoe全局Meta分析AMJHumGenet2016,98(4)680-96
- Horvath S. 脱氧代谢人体组织类型基因组Biol201314(10)R115
- 贝达斯科 MEsteller M.Clinical后天学:抓住翻译机Nat RevGenet201920(2)109-127
- Jones P..!大谷HChakravarthy A德卡瓦略市D. 免疫昆虫学遗传学Nat Rev2019,19(3),151-161
- 阿哈迪S周WSchüssler-Fiorenza Rose, S.M.赛亚尼市R.康波斯K阿维娜M阿什兰市M布鲁内特Snyder,M.个人老化标记和由深纵向剖析显示的老化类型NatMed202026(1)83-90
- 葛拉维娜S东X优Bijg,J.单细胞全双工组测序发现鼠肝类甲片大异性基因组Biol201617(1)150